La plupart du temps, lorsque l’on étudie une population végétale ou animale sur un site, il devient rapidement évident que les individus de cette population ne sont pas toujours répartis de manière uniforme dans l’espace. La dispersion de la population – c'est-à-dire la répartition des individus dans l’espace – est alors d’un intérêt écologique considérable car elle fournit des indications utiles sur l'importance de la compétition intra spécifique, l'aptitude à la dispersion et les systèmes de fécondation.
Non seulement elle conditionne la méthode d’échantillonnage et le choix des techniques d’analyse de données, l’étude de la dispersion constitue une approche incontournable des conditions écologiques de l’habitat et des propriétés biologiques de la population. De plus, tout changement dans le schéma de dispersion doit être analysé parallèlement à des changements de densité dans l’interprétation de la dynamique d’une population. Par exemple, si la dispersion d’une population devient plus régulière, plus uniforme, alors on peut suspecter un accroissement de densité dans la population et une intensification inévitable de la compétition intra spécifique. Une bonne description de la dispersion d’une population est également vitale dans l’analyse des relations proie-prédateur et hôte-parasite modulées par le type de répartition spatiale.
A grande échelle, la population est décrite sur un territoire suffisamment vaste pour que des variations des conditions physico-chimiques entraînent des variations de densité de la population étudiée. On réalise alors une cartographie de la population qui en général peut se superposer à celle des habitats.
A plus petite échelle (en général celle de l’habitat) où les conditions physico-chimiques sont supposées plus homogènes, bien qu’il soit impossible d’en tracer les contours précis, la variabilité aléatoire masque le gradient ou les limites de leur répartition moyenne. En revanche, il est possible de représenter statistiquement le caractère plus ou moins régulier de la densité des organismes étudiés. On parle alors non plus de répartition géographique mais d’un mode de dispersion des organismes dans le milieu, à l’échelle des unités d’observation (quadrat, unité d’effort).
Paramètres de base
Quel que soit l’étude quantitative entreprise, on cherchera à dénombrer les individus soit par comptage exhaustif lorsque c’est possible, mais le plus souvent par échantillonnage. On obtient alors une série de valeurs par unités d’observation (quadrat, unité d’effort), de laquelle on extrait deux paramètres fondamentaux :
1 - La moyenne arithmétique du nombre d’individus par unité d’observation

avec ni = nombre d'unités d'observation présentant la valeur xi et n = nombre total d'observations dans l'échantillon.
2 - La variance du nombre d’individus par unité d’observation
(dans le cas d’un échantillonnage aléatoire simple)

On en déduit les estimations de la moyenne et de la variance de la population :


La précision des estimations de densité dépend de l’effectif de l’échantillon et donc de la variance – paramètre de dispersion autour de la moyenne – qui est sensible à la technique d’échantillonnage employée, elle-même conditionnée par les contraintes du milieu à étudier. L’importance du mode de calcul de la variance n’est pas à négliger car elle intervient de façon décisive dans de nombreux outils de modélisation.
Modèles de distribution
La répartition des individus peut être uniforme. Dans ce cas, on observe à peu près le même nombre d'individus dans chaque unité d’observation et la variance S2 est alors inférieure à la moyenne arithmétique. Au sens biologique, cette distribution implique que le milieu est homogène pour les principales ressources nécessaires à la population et qu'il existe une compétition intra spécifique significative entre les individus. D’un point de vue mathématique, la variable X = "nombre d'individus par unité d’observation" peut être décrite par le modèle binomial B(k,p), avec
k = nombre maximum d'individus par unité d’observation, inconnu et estimé par le nombre maximum d'individus observés dans l'échantillon et
p = µ/k
La probabilité pour que X prenne une valeur r est :

(r = 0, 1, 2, …, k)
La répartition peut aussi être au hasard. Dans ce cas, la variance S2 est proche de la moyenne arithmétique. Sur le plan biologique, ce mode de distribution indique que le milieu est relativement homogène et qu'il n'y a ni compétition ni attraction entre les individus, chacun ayant la même probabilité d’être trouvé en un point donné de l’espace étudié. D’un point de vue mathématique, la variable X = "nombre d'individus par unité d’observation" peut être décrite par le modèle de Poisson P(λ) dans lequel l'espérance est égale à la variance. λ est estimé par la moyenne arithmétique observée dans l'échantillon et
La probabilité pour que X prenne une valeur r est :
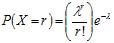
(r = 0, 1, 2, …, k)
Enfin, la répartition peut être en agrégats, encore appelée distribution contagieuse : quelques unités d’observation contiennent beaucoup d'individus, alors que la plupart en possèdent peu ou pas du tout. D’un point de vue écologique, la répartition en agrégats peut résulter de l'hétérogénéité du milieu, de l'attraction que les individus exercent entre eux (chez les insectes très sociaux) ou encore d'une aptitude faible à la dispersion, notamment chez les plantes dont les graines sont dispersées par gravité par des fruits explosifs. D’un point de vue mathématique, la variance est toujours supérieure à la moyenne arithmétique. Un modèle susceptible de décrire cette situation est la loi binomiale négative BN(μ, k), dans laquelle μ représente l'espérance du nombre d'individus par unité d’observation et k mesure la dispersion : plus k est grand, plus la contagion est faible et lorsque k > 5 la répartition devient quasiment aléatoire ; l'inverse de k est donc une mesure de l'agrégation. Ces deux paramètres étant inconnus, μ est estimé par la moyenne arithmétique expérimentale, et k est obtenu au maximum de vraisemblance, en équilibrant la relation :

(pour tout i)
dans laquelle Axi représente le nombre d'observations qui restent à faire lorsqu'on a observé la valeur xi ; l'estimation de k résulte d'un calcul itératif où μ est remplacé par sa valeur expérimentale.
En pratique, une première approximation de k peut être obtenue par

qui donne des résultats satisfaisants la plupart du temps.
La probabilité pour que X prenne une valeur r est :

(r = 0, 1, 2, …)
mais dans la pratique, on peut utiliser la formule de récurrence :

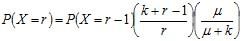
pour r ≥ 1
Ces différentes lois de probabilité permettent un ajustement du modèle théorique candidat aux données expérimentales. Cependant, il est plus prudent au préalable de recourir à une méthode qui permet d'être certain de son choix avec une probabilité d'erreur connue.
Indice de dispersion
De ce qui précède, il apparaît clairement que la relation entre la variance γ et l’espérance μ détermine le modèle de dispersion des individus dans la population. Alors, la rapport

est un indice de dispersion qui prend une valeur inférieure, égale ou supérieure à 1 selon que la répartition des individus est uniforme, au hasard ou en agrégats.
Tester l’hypothèse H0 : "la distribution est au hasard" constitue une méthode rigoureuse qui permet de conclure quant à un type de répartition des individus dans la population. En effet, d'après l'égalité précédente on a :

qui entraine

Par ailleurs, sous l’hypothèse H0, la variable X = "nombre d'individus par unité d’observation" suit une loi de Poisson P(λ) où μ = γ = λ, ce qui permet d'écrire :
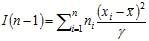
soit
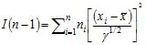
où l’on reconnaît la somme des carrés de n variables centrées réduites indépendantes. Si l'échantillon est suffisamment grand et que λ = γ n’est pas trop petit, ces variables tendent vers une loi normale centrée réduite et I(n-1) suit approximativement une loi de Khi-2 à n-1 degrés de libertés. Dans ces conditions,
On décidera de rejeter l’hypothèse H0 au risque α si :
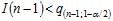
car sous H0, la probabilité de rencontrer une valeur aussi petite est inférieure à α/2.
I est donc jugé inférieur à 1 et la distribution uniforme.
On décidera également de rejeter l’hypothèse H0 au risque α si :

car sous H0, la probabilité de rencontrer une valeur aussi grande est inférieure à α/2.
I est donc jugé supérieur à 1 et la distribution en agrégats.
q(n-1;α/2) étant le quantile de Khi-2 à n-1 degrés de libertés pour un risque d'erreur α.
Une fois le mode de dispersion de l'organisme déterminé, il est toujours intéressant et utile de valider les conclusions par un ajustement du modèle théorique aux données expérimentales.
On peut alors se pencher sur une réflexion relative soit à l'hétérogénéité du milieu ou du niveau de compétition intra spécifique, soit aux conséquences des relations inter spécifiques.
Références
BARBAULT R. (2000). Ecologie générale - Structure et fonctionnement de la biosphère. Dunod, Paris.
FRONTIER S., PICHOD-VIALE D., LEPRÊTRE A., DAVOULT D., LUCZAK C. (2004). Ecosystèmes - Structure, Fonctionnement, Evolution. Dunod, Paris.
HENRY C. (2001). Biologie des populations animales et végétales. Dunod, Paris.
RAMADE F. (2009). Eléments d'écologie - Ecologie fondamentale. Dunod, Paris.
SOUTHWOOD T.R.E., HENDERSON P.A. (2000 - Third edition). Ecological methods. Blackwell Science, Oxford.



 Ancien nom Grec d'une plante utilisée pour teindre les cheveux en blond ; probablement X. strumarium ; de xanthos, jaune, blond - Lampourde à gros fruits, Petite bardane, Glouteron, Herbe aux écrouelles.
Ancien nom Grec d'une plante utilisée pour teindre les cheveux en blond ; probablement X. strumarium ; de xanthos, jaune, blond - Lampourde à gros fruits, Petite bardane, Glouteron, Herbe aux écrouelles. 






















